For instance, knowledge on their behaviour is limited, although herding in sauropodomorphs has been well documented in derived sauropods from the Late Jurassic and Cretaceous.
Here we report an exceptional fossil occurrence from Patagonia that includes over 100 eggs and skeletal specimens of 80 individuals of the early sauropodomorph Mussaurus patagonicus, ranging from embryos to fully-grown adults, with an Early Jurassic age as determined by high-precision U–Pb zircon geochronology.
The presence of sociality in different sauropodomorph lineages suggests a possible Triassic origin of this behaviour, which may have influenced their early success as large terrestrial herbivores.
Soon after dinosaurs originated, early sauropodomorphs (forerunners of the gigantic quadrupedal sauropods) underwent a remarkable adaptive radiation landmarked by the acquisition of herbivory1,2,3,4, large body sizes5, and high taxonomic diversity and specimen abundance2,4,6.Sauropodomorphs were subsequently unaffected by the Triassic–Jurassic extinction event (ca. 200 Ma), which left them as the only large herbivores in terrestrial ecosystems of the Early Jurassic4,6.
The predominance of early sauropodomorph dinosaurs in terrestrial ecosystems extended for almost 40 million years7 (ca. 220–180 Ma, Norian–Pliensbachian8).
Proposed reasons for their early success3,9 include their ability to opportunistically adapt to niches left empty after the extinction of other herbivores1 or to out-perform their competition with superior high-browsing herbivory, large body size, and rapid growth rates4,9,10,11,12,13.
Behaviour has not been regarded as playing a role in the early success of these dinosaurs due to the scarcity of relevant information.
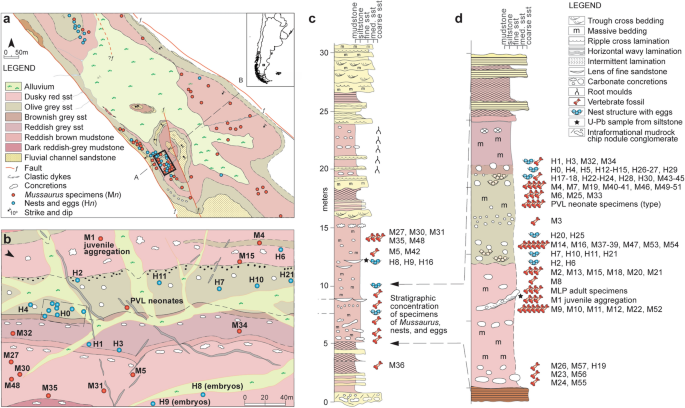
The Laguna Colorada Formation14 of southern Patagonia (Santa Cruz Province, Argentina) contains an exceptional fossil locality (Fig. 1) that provides new information about the social behaviour of early sauropodomorph dinosaurs.The sauropodomorph Mussaurus patagonicus was originally described15 from here based on several well-preserved post-hatchling specimens (Fig. 2) associated with two partially preserved eggs.
(a) general map of fossil findings at the locality (red dots represent skeletal remains of M. patagonicus and blue dots represents eggs or nests, cross section A–B is provided in the Supplementary Information); (b) detailed map of area with high fossil density (including associated juveniles, neonates, and nests), location of this area is indicated with a black rectangle in general map.
Refer to Supplementary Information Fig. 2 showing the structural dip of strata approximately 15 degrees southwest.
patagonicus; (d) detailed stratigraphic section of the 3 m-thick interval with the highest concentration of Mussaurus skeletons and eggs.
Specimens of Mussaurus patagonicus collected from the Laguna Colorada Formation.patagonicus (anterodorsal process of dentary); (e), 3D reconstruction of neonate21 MACN-PV 4111; (f) 3D reconstruction of skull anatomy of neonate MACN-PV 4111; (g), associated skeletons of one-year old juveniles MPM-PV 1813; (h), 3D reconstruction of juvenile MPM-PV 1813/4; (i), associated skeletons of two adult individuals MPM-PV 1868 and MPM-PV 1869; (j), skull of adult individual MPM-PV 1868; (k), Skeletal reconstruction of the different growth stages of M.
patagonicus; (l–o), cortical bone histology of four different sized specimens of M.
patagonicus.
Drawings of silhouettes (by J. Gonzalez) represent the size of the different specimens representing the various ontogenetic stages of M.
patagonicus.
patagonicus specimens (in addition to the 11 individuals discovered decades ago15,17).
In addition, new findings at this site include over 100 eggs in various degrees of association, some of which have preserved embryonic material showing autapomorphic traits of Mussaurus patagonicus (Fig. 2a–c; see Supplementary Information).
All sauropodomorph remains found at this locality have either preserved autapomorphic traits of M.
patagonicus or bear anatomical traits that are indistinguishable from M.
patagonicus, supporting the monospecificity of this unique assemblage.
Covering an area of approximately 1 km2, this rich fossil occurrence is in the type locality of the Laguna Colorada Formation (Fig. 1a–b), which is a 170 m-thick succession of fluvio-lacustrine sediments deposited in a post-rift thermally induced sag basin during the Early Jurassic.All the eggs and specimens of Mussaurus patagonicus were recovered from three distinct horizons within a restricted 3.0 m-thick interval of pedogenically modified massive reddish-brown siltstone in the middle of the Laguna Colorada Formation (Fig. 1c–d).
The latter are interpreted as palustrine carbonate precipitated in massive loessic silts accumulated around a floodplain pond under a seasonally warm climate (see Supplementary Information Fig. 3).
However, we present here U–Pb geochronology (CA-ID-TIMS method) from tuffaceous siltstones intercalated with the vertebrate-bearing interval that yielded two overlapping dates of 192.78 ± 0.14 Ma and 192.74 ± 0.14 Ma (see Supplementary Information).
These dates are younger than previously thought and now assign an Early Jurassic (Sinemurian8) maximum age for the Mussaurus bearing sediments.
Palaeogeographic reconstruction shows the distribution of Late Triassic (purple) and Early Jurassic (blue) of skeletal remains of sauropodomorph dinosaurs (based on fossilworks.org).
Body size graph represents the ancestral reconstruction of the log10BodyMass from the base of Sauropodomorpha to the terminal lineages of sauropods (dark shading represents the minimal values of ancestral reconstruction and light shading the maximal values of ancestral reconstruction).
See supplementary information for details on phylogenetic analysis, optimization of body mass, and phylogenetic diversity of Sauropodomorpha.
Palaeographic map generated with Gplates43 using a Jurassic global palaeogeographic model.
The type specimens of Mussaurus15 originally discovered at this locality consist of eight closely-associated and notably small individuals (estimated body mass = 0.07 kg using volumetric18 and scaling Eqs. 19; see Supplementary Information).Their diminutive body size (Fig. 2d–e) suggests an aggregation of neonates as their body size largely exceeds that of the un-hatched eggs (and the new embryos here reported; Fig. 2c).
The lack of size variation among them suggests they hatched at approximately the same time, and their estimated centre of mass distribution18 and ratio of humeral to femoral shaft circumference20 led to infer a quadrupedal stance for these neonates.
Amongst our recently collected specimens of Mussaurus patagonicus is a remarkable aggregation of at least eleven articulated juvenile skeletons (Fig. 2f–g; estimated body mass = 8.3–10.9 kg using volumetric18 and scaling Eqs. 19; see Supplementary Information), found approximately 50 m from the type hatchlings (Fig. 1b).Each of the bones of these skeletons is surrounded by a 0.5 cm-wide phosphatic halo (Supplementary Information Fig. 12e) which is possibly related to microbial decomposition of soft-tissue22, supporting the interpretation that the skeletons were buried relatively rapidly at the same time (see Supplementary Information).
These histological data combined with the fact that all the specimens are the same size (see Supplementary Information) suggests that these individuals were possibly members of a single brood23 and died together before reaching the first year of life (or, alternatively, they were young juveniles experiencing rapid uninterrupted growth during the first years of life).
Our new collecting also includes nine Mussaurus specimens that are intermediate in size between one-year old juveniles and adults.In the smaller sub-adult specimens (estimated body mass = 106.2 kg using scaling equations 19; see Supplementary Information) a single LAG is usually present, the vascularization is laminar to plexiform, with predominance of circumferentially oriented canals, there are small well-developed primary osteons, and the cortex is formed by fibro-lamellar bone (Fig. 2k).
Larger sub-adult specimens (estimated body mass = 557 kg using scaling equations 19; see Supplementary Information) have over ten LAGs and/or annuli (see Supplementary Information for more detailed histological data, including a characterization of both the LAGs and the annuli).
The largest known specimen of Mussaurus patagonicus (estimated body mass = 1504.8 kg using scaling equations 19; see Supplementary Information) has eight closely spaced LAGs toward the external part of the cortex, which indicates a substantial reduction in growth rate, as preserved on the lateral region of the cross section (Fig. 2m).
Both the estimated centre of mass distribution18 and the ratio of humeral to femoral shaft circumference20 in adult skeletons of Mussaurus unequivocally indicate a bipedal mode of locomotion18,20, contrasting with the inferred quadrupedal stance of neonates (Fig. 2).
patagonicus found in stratigraphically equivalent beds at a nearby locality (see Supplementary Information) were closely associated with each other (Fig. 2h), demonstrating the likelihood of herding behaviour in adult individuals.
The multiple Mussaurus aggregations in the Early Jurassic breeding ground of the Laguna Colorada Formation are interpreted as the oldest skeletal evidence of structured age-segregated gregariousness amongst dinosaurs, pre-dating by over 40 million years reports from Late Jurassic and Cretaceous neosauropods24,25,26.Our new findings on Mussaurus adds important insights on the social behaviour of early sauropodomorphs.
Previous studies reported colonial nesting in the similarly-aged early sauropodomorphs Lufengosaurus from China27 and Massospondylus from South Africa28.In the case of Massospondylus, there also is evidence of site fidelity in its nesting habits28, as in Mussaurus.
The eggs and nests of Mussaurus were found in a sequence of loessic silt, interpreted as windblown dust deposits on the margins of a playa-type arid zone lake.
The similarities in the depositional settings of these localities may suggest an environmental preference by early sauropodomorphs while establishing nesting grounds, although data from other early lineages are needed to test the generality of this hypothesis.
The co-occurrence of associated neonates, juveniles, and adults of Mussaurus in the same restricted stratigraphic interval and at the same locality suggests individuals maintained social cohesion throughout the different stages of their lifespan (notwithstanding possible seasonal variations, as in many modern gregarious species24).In particular, presence of juveniles and sub-adult specimens of Mussaurus at this site would not be expected if only sexually mature individuals congregated temporarily at their preferred nesting area (as in extant crocodiles29).
This is particularly common in extant animals with large body size difference between juveniles and adults, as the activity patterns and foraging habits vary significantly during ontogeny, and the synchronization of these behaviours is crucial for group cohesion30.
The adoption of age-based social partitioning in Mussaurus is compatible with its remarkably large body size increase from hatchlings to adults (0.1 to 1500 kg), the need for several years to attain a subadult body size, and with the different postural habits or locomotion modes (quadrupedal, bipedal) inferred for neonates, juveniles, and adults18.
Mussaurus is deeply nested within the phylogeny of sauropodomorphs17,31,32, and its extensive ghost lineage extends back to the Late Triassic (black arrow in Fig. 3), originating at the large radiation of early sauropodomorphs.The deep origin of the Mussaurus lineage suggests the appearance of structured gregarious behaviour may also extend back to the mid-Norian radiation of sauropodomorphs (at least to the node Sauropodiformes; Fig. 3).However, it is also possible that structured gregarious behaviour may have also arisen at some point during the approximately 18 million years that separate the mid-Norian radiation from the Sinemurian age of Mussaurus.
The massospondylid lineage also originated in the Triassic35 (dark grey arrows in Fig. 3) and the presence of social reproduction in three early sauropodomorph dinosaurs (representing two non-related phylogenetic lineages) allows tracing the appearance of this behaviour back to their most recent common ancestor (close to the node Massopoda; Fig. 3) during the mid-Norian radiation.
Other scenarios require two or more independent origins of social reproduction among sauropodomorphs (e.g., convergently acquired in massospondylids and sauropodiforms).
Finally, the recent discovery of three associated skeletons of the early sauropodomorph Marcocollum in the early Norian of Brazil36 (as well as the Plateosaurus bone-beds from the Norian of Europe) opens the possibility that the origin of some level of gregariousness may extend back to the very base of the Norian radiation (node Plateosauria; light grey arrow in Fig. 3).Further taphonomic information on these assemblages, however, are needed to robustly infer the behavioural implications for early plateosaurian sauropodomorphs, which can be critical for timing the origin of gregariousness in Sauropodomorpha between the early and the mid Norian.
The possible origin of gregariousness in the Norian (in Sauropodiformes, Massopoda, or even Plateosauria) coincides both temporally and phylogenetically with the appearance of evolutionary novelties that have traditionally been linked to the sauropodomorph’s early success as herbivores 3,4,9,11,37.These include multiple anatomical changes in the dentition, skull, and postcranium3 (e.g., neck elongation, reduction of skull size, ventral offset of craniomandibular articulation, overlapping leaf-shaped teeth with coarse obliquely oriented denticles).
However, the most conspicuous change in the early evolution of Sauropodomorpha was the abrupt increase in adult body size5 of over two orders of magnitude (Fig. 3), achieved through the development of accelerated growth rates12,13.
Such a drastic change in body size likely affected multiple aspects of sauropodomorph paleobiology and behaviour, including increased energy requirements and likely larger home ranges and daily foraging distances (both of which are correlated with body size in multiple living species38,39).
Our findings provide important data to temporally link the origin of gregariousness in herbivorous sauropodomorphs to their evolutionary increase in body size.
Social behaviour may represent a previously unrecognised factor that positively influenced the evolutionary path of this clade under a correlated progression model3 (in addition to other anatomical and physiological traits that characterize sauropodomorph body plan).
A key and basic aspect of gregariousness is the synchronization of behaviour and the seasonal environments in which early sauropodomorphs are recorded during the Late Triassic–Early Jurassic (such as the one reported here for Mussaurus) may have influenced the evolution of this features.Early sauropodomorphs are mostly recorded at mid-to-high palaeolatitudes40 during this time (Fig. 3).
Although data on vertebrate assemblages from low paleolatitudes is still scarce, current evidence from well-sampled sequences deposited at low paleolatitudes in the Late Triassic of North America indicates dinosaurs were rare components of the vertebrate fauna and large-bodied sauropodomorphs were absent4,40.
The currently known distribution of early sauropodomorphs show they were predominant and ecologically successful in seasonal environments at mid-to-high palaeolatitudes, as indicated by various paleoclimatic models34,41,42.
Environmental seasonality and the high energetic requirements of large-bodied early sauropodomorphs probably implied long foraging distances (at least during certain periods).
This combination of factors may have favoured the synchronization of behaviour in gregarious sauropodomorphs that required several years to attain subadult body size12.
We postulate the exceptional case of Mussaurus, in which our data show herd behaviour and age-segregation structure, indicates sociality may have influenced the early success of the first global radiation of large-bodied herbivorous dinosaurs.Complete U–Pb data appear in the Supplementary Table 1.
Taxon sampling was expanded adding recently described species from the Late Triassic and Early Jurassic.
Character compilation of these sources resulted in a total of 419 characters (see Supplementary Information).
While the general configuration for PPC-SRµCT was similar for all scans, beam and acquisition parameters were adjusted for each specimen (see Supplementary Information).
All scans were performed using filtered white beam from a wiggler W150B (wiggler gap and filters adjusted per specimen; see Supplementary Table 3).
Images were recorded using an indirect detector consisting of a scintillator (see Supplementary Table 3), a set of optical camera lenses and a PCO.edge 5.5 sCMOS camera with camera link (PCO, Kelheim, Germany).
Pixel size was measured on radiograph, measuring the shift of an object on the sample stage, moved by the most reliable translation motor.
To benefit from the phase aspect of PPC-SRµCT, the sample to detector distance was adjusted considering the energy and the pixel size.
Hence, the exposure time in Supplementary Table 3 is the total integrated time, considering the accumulation of several frames.
Body mass estimates of the different specimens of Mussaurus patagonicus were calculated based on the measurements of femoral circumference using the scaling equation for bipedal non-avian dinosaurs as implemented in the MASSTIMATE 1.3 package for R18, using the quadratic equation implemented in the cQE function with the options equation = raw, cor = 2, and quadratic = TRUE.All data related to the geochronological study, specimen list, and phylogenetic analysis presented in this paper is detailed in the Supplementary Information.Google Scholar .
Testing the effect of the rock record on diversity: A multidisciplinary approach to elucidating the generic richness of sauropodomorph dinosaurs through time.
Google Scholar .
Google Scholar .
Google Scholar .
Cope’s rule and the adaptive landscape of dinosaur body size evolution.
Google Scholar .
Google Scholar .
Extinction of herbivorous dinosaurs linked to Early Jurassic global warming eventL
Google Scholar S
Google Scholar .
The origin and early evolution of dinosaurs.
Google Scholar A
Google Scholar .
An early trend towards gigantism in Triassic sauropodomorph dinosaursA
Google Scholar F
Google Scholar .
Google Scholar .
Skull anatomy of Mussaurus patagonicus (Dinosauria: Sauropodomorpha) from the Late Triassic of Patagonia.
Google Scholar .
Postcranial anatomy and phylogenetic relationships of Mussaurus patagonicus (Dinosauria, Sauropodomorpha).
Google Scholar .
Ontogenetic changes in the body plan of the sauropodomorph dinosaur Mussaurus patagonicus reveal shifts of locomotor stance during growth.
Google Scholar .
Google Scholar .
Google Scholar .
Geochemistry and early diagenesis of mammal-bearing concretions from the sucker creek formation (Miocene) of Southeastern Oregon.
Google Scholar .
Osteohistological insight into the early stages of growth in Mussaurus patagonicus (Dinosauria, Sauropodomorpha)S
Google Scholar .
Google Scholar .
Trackway evidence for a herd of juvenile sauropods from the Late Jurassic of Portugal.
Google Scholar .
Google Scholar .
Embryology of Early Jurassic dinosaur from China with evidence of preserved organic remains.
Google Scholar .
Oldest known dinosaurian nesting site and reproductive biology of the Early Jurassic sauropodomorph Massospondylus.
Google Scholar .
Google Scholar .
Google Scholar .
A new basal sauropodiform from South Africa and the phylogenetic relationships of basal sauropodomorphs.
Google Scholar J
Triassic sauropodomorph dinosaurs from South America: The origin and diversification of dinosaur dominated herbivorous faunas.
Google Scholar .
Google Scholar .
Google Scholar C
Google Scholar .
An exceptionally preserved association of complete dinosaur skeletons reveal the oldest long-necked sauropodomorphs.
Google Scholar .
Early evolution and higher-level phylogeny of sauropod dinosaurs.
Google Scholar .
Dinosaurs, dragons, and dwarfs: The evolution of maximal body size.
Google Scholar .
Home ranges, habitat and body mass: Simple correlates of home range size in ungulates.
Google Scholar .
Google Scholar M
Jurassic climates as inferred from the sedimentary and fossil record.
Google Scholar D
Google Scholar .
Google Scholar J
High-precision U–Pb zircon geochronology of the Late Triassic Chinle Formation, Petrified Forest National Park (Arizona, USA): Temporal constraints on the early evolution of dinosaurs.
Google Scholar .
High-precision U-Pb geochronology and a new chronostratigraphy for the Cañadón Asfalto Basin, Chubut, central Patagonia: Implications form terrestrial faunal and floral evolution in Jurassic.
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar A
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .
Google Scholar .